细胞周期(Cell Cycle)
1953年霍华德(Howard)和培雷克(Pek)率先提出了细胞周期(cell cycle)概念,它构成了细胞学说的重要组成部分。细胞由一次分裂结束到下一次分裂结束,都要经历相同的变化阶段(即G1→S→G2→M )周而复始地进行活动,细胞的这种生长、分裂循环即称为细胞周期(cell cyc1e)。一个细胞周期包括有丝分裂期(M)和分裂间期(G1、S、G2)。尽管在各种细胞中各期所占时间都不尽相同,但相对而言M期最短,S期却较长。
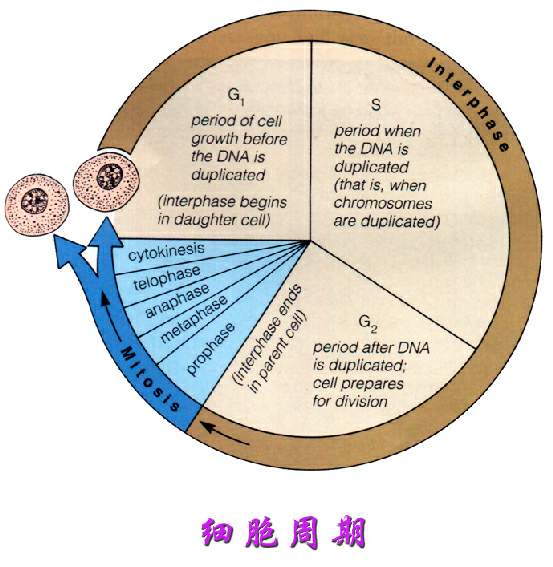
图1为细胞周期
细胞周期(Cell Cycle)是一组有序的事件,最终导致细胞生长和分裂。真核生物的细胞周期可以可分为间隔期和细胞分裂期,间隔期累积有丝分裂所需要的营养素和复制DNA,称为有丝分裂(M)阶段。通过细胞研究的分子机制,分裂期分为三个阶段,G1,S和G2。因此细胞周期包括四个阶段:G1,S,G2和M。
细胞从有丝分裂释放进入 G1 期,此期间 RNA 和蛋白质合成,但没有 DNA 复制。DNA复制的起始标志着 G1 期向 S 期的转化。S 期延伸至所有的 DNA 都被复制完毕。在 S 期,DNA 的全部组成从二倍体 2n 全部复制为 4n。从S期末直至有丝分裂称为G2期,在此期间细胞有两套完整的二倍体染色体。细胞核体积的增加主要在 S 期,此时蛋白质积累,与 DNA 的复制相对应。染色质保持浓缩,形态上没有可见的变化。
细胞周期调节蛋白能与细胞分化周期编码蛋白结合并激活相应的蛋白激酶,从而促进细胞分裂,少发现有11种不同的cyclin,分别为A、B1、B2、C、D1、D2、D3、E、F、G和H。其中8种主要的cyclin己被分离。根据cyclin调控细胞周期时相的不同,可分为G1期和M期两大类。 各类周期蛋白均含有一段约100个氨基酸的保守序列,称为周期蛋白框,介导周期蛋白与CDK结合。动物细胞周期受很多 cdk-周期蛋白复合物控制,激活的CDK1可将靶蛋白磷酸化而产生相应的生理效应,如将核纤层蛋白磷酸化导致核纤层解体、核膜消失,将H1磷酸化导致染色体的凝缩等等。这些效应的最终结果是细胞周期的不断运行。Cdc2 和 cdk 周期蛋白的多种形式被激活的时间说明一种模型,cdk2-G1 周期蛋白二聚体的功能是调控通过 G1 期和 S 期,而 Cdcc2-周期蛋白 A、B 调控有丝分裂过程(Orlando DA,2008)。
已发现很多体内因素可以激发或抑制细胞的增殖,例如多种激素、血清因子、多胺、蛋白水解酶、神经氨酶、cAMP、cGMP以及甘油二脂(DG)、 三磷酸肌醇(IP3)和Ca信使系统等等。细胞内cAMP浓度增加对细胞增殖有抑制作用,凡能使细胞内cAMP增高的因素都能抑制细胞的增殖,降低细胞生长速度;反之,凡能使细胞内cAMP含量下降的因素都能促进DNA的合成与细胞的增殖。细胞周期的各期中的cAMP含量也不相同(见表)。在中国仓鼠卵巢细胞株中,M期cAMP含量最低,M期后cAMP的水平增高三倍,从G1早期至G1晚期,cAMP水平降低到中等水平,直至S期仍维持低的水平。还有许多实验指出cGMP 也对细胞增殖起调控作用,如将cGMP或双丁酰cGMP加到休止在G1期的 3T3 细胞时,能诱导DNA含量的增加, 促进细胞的分裂。如提高细胞cGMP水平,就可促进细胞的有丝分裂,反过来,促进有丝分裂的药物也能增加cGMP的浓度。cAMP能抑制细胞的分裂,促进细胞的分化,cGMP则能抑制细胞分化,促进细胞增殖,在正常生长的细胞中,cAMP和cGMP维持在适当的水平,调节控制细胞周期的运转(Browne G,2010)。