


D-Proline accumulation in plants
Dec 21,2023
General Description
D-Proline accumulation in plants is a common response to various environmental stresses. It serves adaptive functions by acting as a compatible osmolyte, scavenging reactive oxygen species, stabilizing protein structures, regulating cytosolic pH, and influencing stress signaling. The regulation of D-Proline metabolism involves intricate pathways and gene interactions, with the expression of key enzymes being correlated with Pro content. However, externally supplied D-Proline exhibits toxic effects, potentially mediated by its degradation product P5C. Genetic manipulations have shown that enhancing Pro synthesis improves stress tolerance, while impairing Pro catabolism increases sensitivity to D-Proline. Further research is needed to fully understand the mechanisms and contributions of D-Proline and P5C to plant toxicity.

Figure 1. D-Proline
D-Proline accumulation in plants
D-Proline accumulation in plants is a common response to various environmental stresses, including salt, drought, high temperature, low temperature, heavy metal exposure, pathogen infection, anaerobiosis, nutrient deficiency, atmospheric pollution, and UV irradiation. The level of D-Proline accumulation can be significantly higher in stressed plants compared to non-stressed ones, with osmotic stress being the most extensively studied due to its relevance in agriculture. The accumulation of D-Proline is believed to serve adaptive functions in enhancing plant stress tolerance. It has been suggested that D-Proline acts as a compatible osmolyte, serving as a reservoir for carbon and nitrogen. Moreover, under conditions of salinity and drought, which induce oxidative stress, D-Proline has been shown to act as a scavenger for reactive oxygen species (ROS), thereby mitigating oxidative damage. Additionally, D-Proline may function as a molecular chaperone, stabilizing protein structures, while also assisting in regulating cytosolic pH and balancing cellular redox status. Furthermore, D-Proline accumulation may also participate in stress signaling, influencing adaptive responses in plants. The increase in D-Proline accumulation during osmotic stress primarily results from elevated synthesis and reduced degradation within plant cells. Understanding the mechanisms underlying D-Proline accumulation and its adaptive roles in plant stress responses has significant implications for developing stress-tolerant crop varieties and improving agricultural practices. Further research into the regulation and utilization of D-Proline in plants under stress conditions holds promise for enhancing crop resilience and productivity in the face of environmental challenges. 1
Metabolism regulation during development and stress
The regulation of D-Proline metabolism during development and stress in Arabidopsis thaliana involves intricate pathways and gene interactions. In A. thaliana, Pro biosynthesis is controlled by two P5CS isoenzymes, with the rate-limiting step being controlled by feedback inhibition and transcriptional regulation. Additionally, P5CR, encoded by a single gene, is active in both chloroplasts and cytosol. During development, the levels of free Pro vary among plant organs and are dependent on plant age, leaf age, and position. Pro content is correlated with the levels of P5CS and P5CR transcripts and P5CR protein, except in roots, where Pro export via xylem to the shoot plays a role. Under stress, such as osmotic stress, Pro accumulation is induced, with the expression of P5CS being well correlated with Pro content. Different signaling pathways are responsible for the up-regulation of P5CS1 gene during osmotic and salt stress. Furthermore, the regulation of Pro degradation and the importance of P5CDH in Pro accumulation have also been elucidated. This intricate regulatory network involving multiple genes and pathways highlights the complex nature of D-Proline metabolism in A. thaliana, shedding light on its developmental and stress-induced dynamics. 2
Plant toxicity
D-Proline, a derivative of the amino acid L-glutamic acid, plays a crucial role in plant stress responses. However, when externally supplied under normal conditions, D-Proline exhibits toxic effects. The toxicity of D-Proline is a subject of debate, with some studies suggesting that its degradation product, pyrroline-5-carboxylate (P5C), is responsible for the toxicity. Genetic manipulations of Pro metabolic pathways in plants have shed light on its toxicity and impact on osmotolerance. Overexpression of P5CS, the enzyme responsible for Pro synthesis, has been shown to enhance salt tolerance in tobacco plants. Conversely, reduced osmotolerance was observed when the P5CS gene was suppressed or mutated. Overexpression of P5CR, the enzyme involved in D-Proline degradation, did not significantly affect osmotolerance. However, overexpression of P5CR from Arabidopsis in soybean improved drought and heat stress. Studies on Pro catabolism in relation to P5C/GSA (glutamate semialdehyde) accumulation have demonstrated that overexpression of D-Proline dehydrogenase (PDH) decreases sensitivity to externally supplied D-Proline, while decreased PDH activity or knockout mutations increase sensitivity to Pro. Furthermore, external application of P5C has been shown to induce cell death, increase reactive oxygen species production, and inhibit plant growth. While there is evidence supporting P5C as a mediator of Pro toxicity, other data suggests that Pro itself can induce toxicity. High external concentrations of D-Proline have been found to be toxic to Arabidopsis plants, and mutants with impaired D-Proline catabolism display increased sensitivity to D-Proline. Transcriptomic analysis of these mutants treated with D-Proline revealed the repression of genes related to photosynthesis and cell-wall protein synthesis, which are believed to contribute to D-Proline toxicity. In summary, the toxicity of D-Proline in plants is a complex phenomenon involving both D-Proline and its degradation product P5C. Further research is needed to fully understand the underlying mechanisms and clarify the relative contributions of D-Proline and P5C to plant toxicity. 3
Reference
1. Verbruggen N, Hermans C. Proline accumulation in plants: a review. Amino Acids. 2008 Nov;35(4):753-759.
2. Verbruggen N, Hua XJ, May M, Van Montagu M. Environmental and developmental signals modulate proline homeostasis: evidence for a negative transcriptional regulator. Proc Natl Acad Sci USA, 1996, 93:8787–8791.
3. Ayliffe MA, Roberts JK, Mitchell HJ, Zhang R, Lawrence GJ, Ellis JG, Pryor JT. A plant gene up-regulated at rust infection sites. Plant Physiol, 2002, 129:169–180.
- Related articles
- Related Qustion
- D-Proline: Biological Distribution and Applications in Pharmaceutical Synthesis Apr 30, 2024
D-Proline, vital in pharmaceutical synthesis, aids in drug creation, especially in asymmetric synthesis and peptide-based development, advancing pharmaceutical research.
- D-proline:A member of the D-amino acid family Sep 19, 2023
D-proline(D-Pro), also known as D-hydroxypyrrole carboxylic acid, is an important five-membered cyclic amino acid and an important chiral compound.
- Split preparation D-proline Nov 5, 2021
The price ratio of D-proline and L-proline is quite different. Therefore, D-proline is prepared from L-proline through asymmetric transformation. Proline has a very important meaning.

Supplementation with pyridoxal 5'-phosphate monohydrate can synthesize neurotransmitters such as dopamine and serotonin, maintaining a healthy nervous system.....
Nov 4,2025Biochemical Engineering
Remdesivir is synthesised using ribonolactone as a raw material through reactions in the presence of a pre-drug coupling partner (P-Stereogenic Prodrug Coupling Partner).....
Dec 21,2023Inhibitors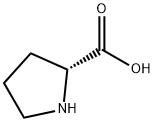
D-Proline
344-25-2You may like
- D-Proline
-
- $0.00 / 1kg
- 2025-12-12
- CAS:344-25-2
- Min. Order: 1kg
- Purity: 98.5%-101%
- Supply Ability: 500kg
- (R)-pyrrolidine-2-carboxylic acid
-
- $29.00 / 1mL
- 2025-12-11
- CAS:344-25-2
- Min. Order:
- Purity: 99.91%
- Supply Ability: 10g
- D-Proline
-

- $0.00 / 1kg
- 2025-12-10
- CAS:344-25-2
- Min. Order: 1kg
- Purity: 98%
- Supply Ability: 20t