背景及概述[1][2]
寡霉素抗生素组合物是由美国威斯康辛大学的Smith等在1954年首次从淀粉酶产色链霉菌(Streptomyces diastatochromogenes)中分离得到的一类结构相似的二十六元环大环内酯类抗生素,包括3个不同比例的主要组分A、B和C,这3个主要组分的比例取决于微生物生产菌株和培养条件它们化学结构的区别在于C125~C128位所连接的基团不同。
其生物学活性多种多样,包括抗真菌活性、抗肿瘤活性、呼吸抑制作用及杀虫活性等。虽然它们在临床上尚未被应用,但作为ATP合酶抑制剂,寡霉素具有重要的科学意义,已被广泛应用于科学研究当中。它们早已作为化学试剂被销售,价格昂贵,具有一定经济价值。
生物学活性及应用[1][2]
1. 抗真菌活性
寡霉素具有广谱的抗真菌活性。例如寡霉素ABC混合物可强烈抑制黑曲霉(Aspergillus niger),多孔木霉(Tolypocladium inflatum),新月弯孢菌(Curvularialunata),Fusarium ocsispoFum和Trichoderma alba,尤其对人类致病菌皮炎芽生菌(Blastomyees dermatitidis)具有显著的抑制活性。
一般地,寡霉素A、B和C的抗真菌活性强度为A>B>C。寡霉素A对植物致病菌灰葡萄孢菌(Botrytis cinerea),黄瓜黑星病菌(Cladosporium cucumerinum),黄瓜炭疽病菌(Colletotrichum lagenarium),稻瘟病菌(Magnaporthegrisea)和南瓜疫病菌(Phytophthora capsici)的最低抑制浓度(MIC)为3~5ktg/mL。
2. 抗肿瘤活性
寡霉素A、B和C具有极强的抗肿瘤活性。在以线粒体作为靶标的37,000个化合物对人类60个肿瘤细胞株的抗癌活性试验中,寡霉素是效果的37种化合物之一。抗阿霉素的肿瘤细胞株R.HepG2细胞可产生P-糖蛋白,积累的阿霉素比亲代细胞少。寡霉素可阻止P-糖蛋白的活性,使R-HepG2细胞积累更多阿霉素,从而引发细胞的程序性死亡。
2003年发现,寡霉素ABC的混合物在浓度为3pg/mL时开始抑制小鼠P388淋巴性白血病细胞的存活,在30pg/mL时使细胞存活率下降到54%,但是随着寡霉素ABC的浓度进一步加大,细胞存活率不再下降。这些浓度比完全抑制各种细胞呼吸作用所需浓度(80~300nmol/L)要低1 000倍以上。核损伤、染色体浓缩和破碎是程序性死亡的标志。
寡霉素ABC在1 0pg/mL时抑制细胞从G1期到s期的转变,但并未引起DNA降解(因subG1期细胞的比例保持不变)。在浓度为30pg/mL时,含有亚二倍体DNA的细胞比例比对照试验增加3倍。损伤的核的比例随着寡霉素ABC浓度的增加而增加。试验结果表明,寡霉素ABC在浓度超过10pg/mL时可诱导P388细胞的程序性死亡(因为含亚二倍体DNA的细胞比例的增加是程序性死亡的一个征兆),抑制细胞分裂并诱导细胞死亡是导致细胞存活率下降的原因。
寡霉素A对P388细胞的生长抑制作用与寡霉素ABC的混合物类似,但是抑制浓度更低,即寡霉素A在lpg/mL时就开始抑制细胞的存活,在10pg/mL时达到抑制率。实验结果表明,50pg/mL寡霉素A对人类肿瘤细胞Be1.7402、K.562和HCT-8的增殖抑制率分别为29.9%,43.7%和53.5%。而在腹(膜)腔内给药的方式下,寡霉素A、B和C对小鼠的半致死量(LD )分别为1.5、2.9和8.3mg~gt。因此,寡霉素作为抗肿瘤剂极具应用潜力。
3. 氧化磷酸化抑制作用
寡霉素是哺乳动物细胞氧化磷酸化的抑制剂。它能有效地结合线粒体FnF ATP合酶的功能亚基F ,使ATP合酶的构型发生改变,从而抑制线粒体膜间隙的质子流回流到线粒体基质,其结果是ATP的合成被阻断,造成生物代谢所需能量不足,因此寡霉素对哺乳动物具有很强的毒性。寡霉素ABC混合物完全抑制各种细胞呼吸作用所需浓度为80-300nmol/L(按寡霉素ABc混合物的平均分子量为791计算,则相当于0.0632~0.237~tg/mL,寡霉素A完全抑制一些肿瘤细胞呼吸作用所需浓度仅为100ng/mL。
作为ATP合酶抑制剂,寡霉具有重要的科学意义。将寡霉素应用到科研实验工作的先锋人物,这为后来氧化磷酸化过程的阐明做出了巨大贡献。细胞生物能量的改变与许多疾病过程有关,对使真核细胞产生大部分ATP的酶一F1F0.ATP合酶进行调节,有望用于这些疾病的治疗。线粒体是程序性死亡的关键性调节因素,这意味着线粒体可作为癌症治疗靶标。因此,为线粒体氧化磷酸化抑制剂的寡霉素,被广泛应用于氧化磷酸化、与线粒体功能紊乱有关疾病、及程序性死亡等相关研究。
除以上几种主要生物学活性外,寡霉素A、B和C还具有杀虫活性,可用于杀死蚂蚁、甲虫和线虫等昆虫。
制备[1]
寡霉素可由多种链霉菌产生。而由多种链霉菌产生的次级代谢物的基因簇,通常定位于6.5Mb的染色体内部保守区域。例如在阿维链霉菌中,土味素(位点2,635,583~2,640,003)、戊丙酯菌素;位点3,749,307~3,758,093)和寡霉素(位点3,534,525~3,634,592)的生物合成基因簇都定位于6.5Mb的内部区域。寡霉素属于I型聚酮(polyketide)化合物,其生物合成基因簇在阿维链霉菌9个I型聚酮合酶(PKS)基因簇中。根据相关网站公布的最新数据,寡霉素生物合成基因簇位点为3,534,525~3,634,592bp,全长约100kb,共有14个开放阅读框架。
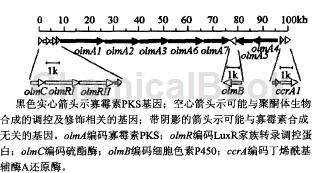
在基因簇内部长约89.6kb的片段含有7个大的开放阅读框架(olmA1.olmA2.olmA3.olmA6-olmA7)和(olmA5-olmA4),其中olmA 1、olmA2、olmA3、olmA6和olmA7的转录方向相同,D,m 5和0 4的转录方向相同。这7个基因共同编码多功能的PKS。该PKS由17个模块组成(其中包括一个装载模块),共有79个催化酶域,但其中一些催化酶域可能是非功能性的。
通过转座对olmA4区域进行破坏,获得不产寡霉素的突变株,表明olmA4参与寡霉素的生物合成。在olmA 1的上游是寡霉素生物合成基因簇的调控基因olmR I~olmR II,根据生物信息学分析推测它们编码LuxR家族的转录调控蛋白。推测在olmR I上游是编码硫酯酶(thioesterase1的基因olmC,聚酮可能在该硫酯酶的作用下从PKS上释放出来;在olmA7和olmA5之间是编码细胞色素P450羟化酶的基因olmB;基因簇最右端是编码丁烯酰基辅酶A还原酶的基因ccrA1;olmRII与olmA1之间的一个开放阅读框架可能是个假基因;而olmA1与ccrA1之间的一个开放阅读框架的推测功能未知。
除了编码PKS、转录调控蛋白和硫酯酶的基因外,其他的一些基因可能与聚酮体的修饰有关。另外,最近的研究表明,在阿维链霉菌中,阿维菌素的途径特异性正调控蛋白AveR对于寡霉素的产生具有负调控作用。寡霉素和阿维菌素都是I型聚酮化合物。以前的研究表明,寡霉素与阿维菌素具有相同的生物合成前体(如乙酸盐和丙酸盐)。
主要参考资料
[1] 寡霉素的研究进展
[2] 寡霉素对白血病K562/A02细胞株凋亡调控机制探讨