


BOC-L-Leucine: Synthesis, Role in Glucose Homeostasis and Potential Risks
May 7,2025
BOC-L-Leucine is an essential amino acid for protein synthesis. Additionally, similarly to other amino acids, the carbon skeleton of leucine can be used to generate ATP. However, leucine can also regulate several cellular processes such as protein synthesis, tissue regeneration, and metabolism. Therefore, leucine supplementation has been studied in a variety of conditions such as aging, muscle lesions, protein/energy deprivation, obesity, and diabetes mellitus.

Synthesis of BOC-L-Leucine
General procedure: Boc-protected amino acids were prepared following a literature method. Initially, to a solution of water/1,4-dioxane (3:2 v/v), 5.0mmol of an amino acid (l-valine, l-leucine, l-phenylalanine or l-proline) and 0.3g of NaHCO3 were added. Next, 7.5mmol of Boc anhydride (1.5 equiv.) were added to this mixture. The reaction mixture was stirred at room temperature for 24h. The crude reaction containing BOC-L-Leucine mixture was then diluted in water (50.0mL) and a liquid-liquid extraction employing 40mL of ethyl acetate was carried out to remove the residual Boc2O. The pH of the water phase was then adjusted to 1 with an aqueous solution of HCl (2M). Next, a liquid-liquid extraction employing ethyl acetate was carried out (three aliquots of 100.0mL). The organic phase was then dried under reduced pressure and further purification by column chromatography (silica gel) employing isocratic eluent 9:1 dichloromethane/methanol was carried out. The solvent was removed under reduced pressure to provide BOC-L-Leucine which was used directly for next reaction without further purification (yield, 97%). 1H NMR (400 MHz, CDCl3) δ 4.89 (broad d, 1H, J=8.3 Hz), 4.31 (m, 1H), 1.74-1.49 (m, 3H), 1.44 (s, 9H), 0.95 (d, 6H, J=6.5 Hz).[1]
A 50 mL three-necked flask equipped with a thermometer, a constant-pressure titration funnel and a glass stopper was charged with the compound L-The acid V (5.0 g, 0.038g NaOH (3.0 g, 0.075g Mmol) and 15 mL of water were added, Stirring vigorously for 5 min,So that the material dissolved completely.(Boc) 20 (10.8g, 10g) was slowly added dropwise at room temperature in a water bath at 25 to 30 ° C; (50 mol) in THF (5 mL)After the dropwise addition, slowly raise the temperature to 50 ~ 55 ° C.After 5h, NaOH (1.5g, 0.038m) was added to the system,In water (5 mL)Insulation reaction 3h,After TLC detection reaction was complete,Ethyl acetate (20 mL) was added to the system.Ice water bath,The pH was adjusted to 2 to 3 with 6N dilute hydrochloric acid, the ethyl acetate phase was filtered,The aqueous phase with BOC-L-Leucine was washed with ethyl acetate (20 mL)The ethyl acetate phase was combined,Followed by saturated NaHCO3,The organic phase was washed with saturated NaCl,Na2SO4 was added to dry the organic phase,And the organic phase was distilled off with a rotary evaporator,The crude product 8. 82g.Recrystallization from n-hexane (10 mL), filtration,The filter cake was dried under reduced pressure,To be fine BOC-L-Leucine 47.61g, yield 86.7%.[2]
Regulation of Glucose Homeostasis by BOC-L-Leucine
Previous studies observed improved glucose tolerance in leucine-treated animals. For example, BOC-L-Leucine supplementation in the drinking water prevented HFD-induced hyperglycemia and insulin resistance in mice. Although a reduced fat mass may explain a portion of these effects, Eller et al. observed that the improved glucose control occurred independently of changes in body composition. Therefore, leucine supplementation may improve glucose homeostasis and prevent at least part of diet-induced insulin resistance. The underlying mechanisms involved in the effects of leucine on glucose control remain unknown. The pancreas is a potentially important leucine-target tissue that might affect glucose homeostasis. BOC-L-Leucine stimulates protein synthesis in pancreatic β and acinar cells through the mTOR signaling pathway. Additionally, leucine presents insulinotropic properties. Therefore, increased insulin secretion in leucine-supplemented individuals could improve their postprandial glucose levels. However, some caution when using leucine supplementation in specific situations is recommended. BOC-L-Leucine supplementation in pregnant rats resulted in decreased β-cell formation in their offspring, which could potentially increase the risk of type 2 diabetes mellitus later in life. Additionally, leucine supplementation enhances tumor growth in a murine model of pancreatic cancer.[3]
Other mechanisms, in addition to the regulation of insulin secretion, are likely to be involved in the effects of BOC-L-Leucine on glucose control. Several studies have observed that leucine-treated animals exhibited improved insulin sensitivity. Additionally, BOC-L-Leucine supplementation decreases glucose-6-phosphatase expression in the livers of mice consuming an HFD, suggesting reduced gluconeogenesis. Leucine supplementation increases SIRT1 expression and prevents mitochondrial dysfunction in the livers of diet-induced obese mice. Furthermore, hepatic steatosis and lipid metabolism were improved in leucine-supplemented animals. BCAA or leucine supplementation also affects glucose metabolism and glycogen synthesis in skeletal muscle. Therefore, the direct effects of BOC-L-Leucine on insulin-sensitive tissues such as the liver and skeletal muscle may influence whole-body glucose homeostasis. Paradoxically, classical studies have found that high physiological BCAA concentrations inhibit the early steps in insulin signaling and that amino acid infusion acutely causes insulin resistance in human skeletal muscle. More recently, some studies revealed that obese and lean humans differ in terms of BCAA metabolism and that BCAA may contribute to insulin resistance in obesity in humans. Balage et al. observed that five-week BOC-L-Leucine supplementation induced a delay in postprandial stimulation in the early steps of muscle insulin signaling, leading to overall glucose intolerance. Insulin resistance is also induced by mTOR/S6K1 pathway overactivation. Accordingly, the absence of S6K1 protects against age- and diet-induced obesity and enhances insulin sensitivity. Additionally, hyperinsulinemia leads to insulin resistance in the liver and skeletal muscle through a rapamycin-sensitive mechanism. Because adipose tissue is an important leucine-responsive tissue, leucine supplementation may change the secretion pattern of adipokines to a more favorable profile. Previous studies have found that BOC-L-Leucine supplementation in previously obese rats increased adiponectin levels. In another study, leucine supplementation reduced inflammatory marker levels in white adipose tissue.
References
[1]Martins, Rafaela de S.; Pereira, Mathias P.; de Castro, Pedro P.; Bombonato, Fernanda I.[Tetrahedron, 2020, vol. 76, # 5, art. no. 130855
[2]SHANGHAI INSTITUTE OF TECHNOLOGY - CN105294501, 2016, A
[3]Pedroso JA, Zampieri TT, Donato J Jr. Reviewing the Effects of L-Leucine Supplementation in the Regulation of Food Intake, Energy Balance, and Glucose Homeostasis. Nutrients. 2015 May 22;7(5):3914-37.
- Related articles
- Related Qustion

Supplementation with pyridoxal 5'-phosphate monohydrate can synthesize neurotransmitters such as dopamine and serotonin, maintaining a healthy nervous system.....
Nov 4,2025Biochemical Engineering
In molecular biology, Diethyl pyrocarbonate (DEPC) is mainly used as a strong, but not absolute, inhibitor of RNase activity.....
May 7,2025Inhibitors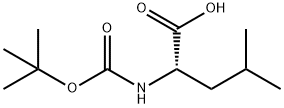
BOC-L-Leucine
13139-15-6You may like
- BOC-L-Leucine
-
- $0.00/ kg
- 2025-12-12
- CAS:13139-15-6
- Min. Order: 1kg
- Purity: 98%
- Supply Ability: 1T+
- BOC-L-Leucine
-

- $0.00 / 25Kg/Drum
- 2025-12-12
- CAS:13139-15-6
- Min. Order: 1KG
- Purity: 98%min
- Supply Ability: 500kgs
- BOC-L-Leucine
-

- $10.00 / 1KG
- 2025-12-11
- CAS:13139-15-6
- Min. Order: 1KG
- Purity: 99%
- Supply Ability: 5tons