


Potential pharmacological effects of L-Arabinose
Sep 10,2025
Introduction
L-Arabinose (L-Ara) is a plant saccharide that is not found in animals. Like xylose, L-Arabinose is a pentose comprising five carbons, not six carbon containing hexose like glucose (Glc) and galactose (Gal)(Figure 1). Although its content in the cell walls varies depending on the plant species, L-Arabinose can be considered a major sugar. It accounts for 5-10 % of cell wall sugar, for instance, in Arabidopsis (Arabidopsis thaliana) and rice (Oryza sativa). The fact that L-Arabinose is widely distributed not only in land plants including liverworts and mosses but also found in several Chlorophycean and Charophycean green algae suggests that the metabolic pathway for the synthesis of L-Arabinose was acquired early by primitive plants. The broad range of L-Arabinose-containing molecules seen in land plants today is likely due to subsequent diversification of the use of L-Arabinose during plant evolution.

L-Arabinose may be useful as a natural pharmaceutical. Monomeric L-Arabinose inhibits intestinal maltase and sucrase(α-glucosidase hydrolyzing sucrose) activities in vitro. In rats, dietary sucrose increases the insulin level in blood and triacylglycerol levels in blood plasma and the liver, but feeding L-Arabinose together with sucrose can significantly reduce the increase in these levels. Recently, the effect of L-Arabinose on controlling insulin and blood-Glc levels was also observed in humans. While its effect in humans is still controversial, the use of l-Ara for these purposes is receiving attention and becoming more wide-spread.[1]
L-Arabinose promotes expansion in superspreaders
The molecular understanding of host-pathogen interactions in the gastrointestinal (GI) tract of superspreader hosts is incomplete. In a mouse model of chronic, asymptomatic Salmonella enterica serovar Typhimurium (S. Tm) infection, we performed untargeted metabolomics on the feces of mice and found that superspreader hosts possess distinct metabolic signatures compared with non-superspreaders, including differential levels of L-arabinose. RNA-seq on S. Tm from superspreader fecal samples showed increased expression of the L-arabinose catabolism pathway in vivo. By combining bacterial genetics and diet manipulation, we demonstrate that diet-derived L-arabinose provides S. Tm a competitive advantage in the GI tract, and expansion of S. Tm in the GI tract requires an alpha-N-arabinofuranosidase that liberates L-arabinose from dietary polysaccharides. Ultimately, our work shows that pathogen-liberated L-arabinose from the diet provides a competitive advantage to S. Tm in vivo. These findings propose L-arabinose as a critical driver of S. Tm expansion in the GI tracts of superspreader hosts.[2]
L-Arabinose transport and Metabolism in Salmonella Influences Biofilm Formation
L-arabinose inducible promoters are commonly used in gene expression analysis. However, nutrient source and availability also play a role in biofilm formation; therefore, L-arabinose metabolism could impact biofilm development. In this study we examined the impact of L-arabinose on Salmonella enterica serovar Typhimurium (S. Typhimurium) biofilm formation. Using mutants impaired for the transport and metabolism of L-arabinose, we showed that L-arabinose metabolism negatively impacts S. Typhimurium biofilm formation in vitro. When L-arabinose metabolism is abrogated, biofilm formation returned to baseline levels. However, without the ability to import extracellular L-arabinose, biofilm formation significantly increased. Using RNA-Seq we identified several gene families involved in these different phenotypes including curli expression, amino acid synthesis, and L-arabinose metabolism. Several individual candidate genes were tested for their involvement in the L-arabinose-mediated biofilm phenotypes, but most played no significant role. Interestingly, in the presence of L-arabinose the diguanylate cyclase gene adrA was downregulated in wild type S. Typhimurium. Meanwhile cyaA, encoding an adenylate cyclase, was downregulated in an L-arabinose transport mutant. Using an IPTG-inducible plasmid to deplete c-di-GMP via vieA expression, we were able to abolish the increased biofilm phenotype seen in the transport mutant. However, the mechanism by which the L-arabinose import mutant forms significantly larger biofilms remains to be determined. Regardless, these data suggest that L-arabinose metabolism influences intracellular c-di-GMP levels and therefore biofilm formation. These findings are important when considering the use of an L-arabinose inducible promoter in biofilm conditions.[3]
L-Arabinose affects the c-di-GMP and global gene expression of Vibrio parahaemolyticus.
The L-arabinose inducible pBAD vectors are commonly used to turn on and off the expression of specific genes in bacteria. The utilization of certain carbohydrates can influence bacterial growth, virulence factor production, and biofilm formation. Vibrio parahaemolyticus, the causative agent of seafood-associated gastroenteritis, can grow in media with L-arabinose as the sole carbon source. However, the effects of L-arabinose on V. parahaemolyticus physiology have not been investigated. In this study, we show that the growth rate, biofilm formation capacity, capsular polysaccharide production, motility, and c-di-GMP production of V. parahaemolyticus are negatively affected by L-arabinose. RNA-seq data revealed significant changes in the expression levels of 752 genes, accounting for approximately 15.6% of V. parahaemolyticus genes in the presence of L-arabinose. The affected genes included those associated with L-arabinose utilization, major virulence genes, known key biofilm-related genes, and numerous regulatory genes. In the majority of type III secretion system, two genes were upregulated in the presence of L-arabinose, whereas in those of type VI secretion system, two genes were downregulated. Ten putative c-di-GMP metabolism-associated genes were also significantly differentially expressed, which may account for the reduced c-di-GMP levels in the presence of L-arabinose. Most importantly, almost 40 putative regulators were significantly differentially expressed due to the induction by L-arabinose, indicating that the utilization of L-arabinose is strictly regulated by regulatory networks in V. parahaemolyticus. The findings increase the understanding of how L-arabinose affects the physiology of V. parahaemolyticus. Researchers should use caution when considering the use of L-arabinose inducible pBAD vectors in V. parahaemolyticus. IMPORTANCE The data in this study show that L-arabinose negatively affects the growth rate, biofilm formation, capsular polysaccharide production, motility, and c-di-GMP production of V. parahaemolyticus. The data also clarify the gene expression profiles of the bacterium in the presence of L-arabinose. Significantly differentially expressed genes in response to L-arabinose were involved in multiple cellular pathways, including L-arabinose utilization, virulence factor production, biofilm formation, motility, adaptation, and regulation. The collective findings indicate the significant impact of L-arabinose on the physiology of V. parahaemolyticus. There may be similar effects on other species of bacteria. Necessary controls should be established when pBAD vectors must be used for ectopic gene expression.[4]
L-Arabinose suppresses gluconeogenesis
L-Arabinose is a kind of plant-specific five-carbon aldose with benefits in type 2 diabetes mellitus. It has been shown to have good properties in improving glucose homeostasis, but the underlying molecular mechanisms are still not clear. Hepatic gluconeogenesis is critical for regulating glucose homeostasis. Here, this study aimed to investigate whether l-arabinose could improve glucose metabolism via suppressing hepatic gluconeogenesis. High-fat-high-sucrose diet (HFHSD) or high-sucrose diet (HSD)-fed mice were supplemented with or without l-arabinose for 12 weeks. Fasting blood glucose levels were measured and glucose tolerance test and the histological analysis were performed after l-arabinose administration. AMP-activated protein kinase (AMPK), acetyl-CoA carboxylase (ACC), peroxisome proliferator activated receptor-γ coactivator-1α (PGC1α), Forkhead box O1 (FoxO1), phosphoenolpyruvate carboxykinase (PEPCK) and glucose-6-phosphatase (G6Pase) expression levels were determined by RT-PCR and western blotting. As expected, l-arabinose apparently decreased body weight and attenuated hyperglycemia and glucose intolerance caused by HFHSD or HSD. l-Arabinose also had beneficial effects on glycogen synthesis by inactivating GSK3β. The expression levels of gluconeogenic genes were all decreased by l-arabinose administration in vivo and in vitro. In addition, our work revealed that AMPK is required for the inhibitory effects of l-arabinose on hepatic gluconeogenesis. l-Arabinose significantly up-regulated the phosphorylated levels of AMPK and its downstream protein ACC. Furthermore, blocking AMPK signaling through an inhibitor (compound C) or siAMPK significantly attenuated the inhibition of hepatic gluconeogenesis and the promotion of glycogen synthesis with l-arabinose, indicating that the inhibitory effect of l-arabinose on hepatic gluconeogenesis was AMPK dependent. These work revealed that l-arabinose is a promising natural product for the regulation of hyperglycemia through inhibition of hepatic gluconeogenesis by activating AMPK.[5]
References
[1]Kotake T, Yamanashi Y, Imaizumi C, Tsumuraya Y. Metabolism of L-arabinose in plants. J Plant Res. 2016;129(5):781-792. doi:10.1007/s10265-016-0834-z
[2]Ruddle SJ, Massis LM, Cutter AC, Monack DM. Salmonella-liberated dietary L-arabinose promotes expansion in superspreaders. Cell Host Microbe. 2023;31(3):405-417.e5. doi:10.1016/j.chom.2023.01.017
[3]Vasicek EM, O'Neal L, Parsek MR, Fitch J, White P, Gunn JS. L-Arabinose Transport and Metabolism in Salmonella Influences Biofilm Formation. Front Cell Infect Microbiol. 2021;11:698146. Published 2021 Jul 22. doi:10.3389/fcimb.2021.698146
[4]Zhang M, Luo X, Li X, et al. L-arabinose affects the growth, biofilm formation, motility, c-di-GMP metabolism, and global gene expression of Vibrio parahaemolyticus. J Bacteriol. 2023;205(9):e0010023. doi:10.1128/jb.00100-23
[5]Wang Y, Guan Y, Xue L, et al. L-Arabinose suppresses gluconeogenesis through modulating AMP-activated protein kinase in metabolic disorder mice. Food Funct. 2021;12(4):1745-1756. doi:10.1039/d0fo02163f
- Related articles
- Related Qustion

Supplementation with pyridoxal 5'-phosphate monohydrate can synthesize neurotransmitters such as dopamine and serotonin, maintaining a healthy nervous system.....
Nov 4,2025Biochemical Engineering
4-Methoxybenzylamine plays an important role in organic synthesis and is often used in the preparation of pharmaceutical intermediates and fine chemicals.....
Sep 10,2025Pharmaceutical intermediates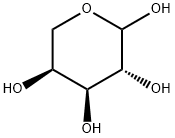
L-Arabinose
87-72-9You may like
- L-Arabinose
-

- $0.00 / 1kg
- 2025-12-22
- CAS:87-72-9
- Min. Order: 1kg
- Purity: 99%
- Supply Ability: 20MT
- L-Arabinopyranose
-
- $72.00 / 100mg
- 2025-12-20
- CAS:87-72-9
- Min. Order:
- Purity: 99.86%
- Supply Ability: 10g
- L-Arabinopyranose
-
- $72.00 / 100mg
- 2025-12-20
- CAS:87-72-9
- Min. Order:
- Purity: 99.86%
- Supply Ability: 10g