摘要
椰子油的主要脂肪酸是月桂酸,约占45-53%。月桂酸对椰子油的许多特性起着重要作用。椰子油在体内很容易被我们消化吸收,月桂酸也很容易被运输,所以代谢很快。详细的研究表明,摄入的大部分月桂酸直接输送到肝脏,在那里,月桂酸直接转化为能量和其他代谢物,而不是作为脂肪储存。这种代谢物包括酮体,可以被肝外组织,如大脑和心脏,作为一种直接的能量形式使用。月桂酸对血清胆固醇影响的研究是矛盾的。在饱和脂肪酸中,月桂酸对脂肪积累的贡献最小。月桂酸和单月桂酸甘油酯对革兰氏阳性菌、多种真菌和病毒具有明显的抗菌活性。今天有许多商业产品使用月桂酸和单月桂酸甘油酯作为抗菌剂。由于月桂酸与长链脂肪酸在性质上的显著差异,它们通常被区分为C6–C12的中链脂肪酸和C14及以上的长链脂肪酸。
符号和缩写

简介
椰子油是存在于椰肉中并从椰肉中提取的食用油。它的独特之处在于它以月桂酸(C12)为主要脂肪酸,占总脂肪酸组成的45-53%。因为椰子油在热带地区被广泛食用,所以在这些地区,[1]月桂酸是饮食的重要组成部分。
虽然植物油的性质不能完全由单一脂肪酸成分来解释,但本综述旨在表明椰子油的许多性质确实可以归因于月桂酸的性质。特别是,本文将讨论月桂酸在中链甘油三酯代谢中的作用,并着重讨论月桂酸本身的代谢。还将讨论月桂酸在胆固醇、脂肪积累和储存等健康问题中的作用。最后,我们将详细介绍月桂酸和单月桂酸甘油酯的抗菌活性。在这些讨论中,一个常见的观察结果是,月桂酸,一种中链脂肪酸,有许多特性,使其区别于长链脂肪酸。
中链脂肪酸(MCFA)和中链甘油三酯(MCT)。“中链脂肪酸”(MCFA)一词尚无公认的定义。该术语通常用于指两种脂肪酸组:C8和C10以及C6–C12。类似地,术语中链甘油三酯(MCT)用于指两种主要类型的甘油三酯:一种仅含有C8和C10脂肪酸的市售甘油三酯混合物,另一种是主要由C6–C12脂肪酸组成的甘油三酯混合物(图1)。
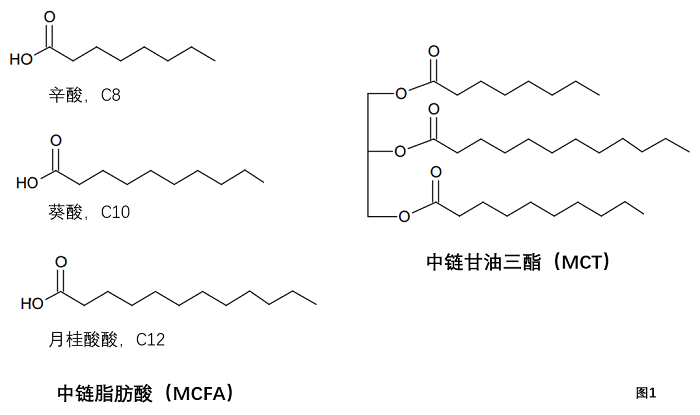
图1. 中链脂肪酸(MCFA)和中链甘油三酯(MCT)的示例
MCT的定义是C8和C10,是这些脂肪酸是从水解椰子油的分馏工业中获得的,用于制造只含有C8和C10的特殊膳食油,而更具商业价值的C12则用于表面活性剂工业[2]。1968年,Harkins和Sarett [3]开发了一种由75%C8和25%C10组成的合成甘油三酯混合物,称为“MCT油”[4],与玉米油等长链脂肪酸(LCFA)植物油相比,MCT油更容易被消化和吸收。MCT油可用于脂肪消化不良患者的营养管理。除了在临床上使用外,现在运动员和医学研究人员还使用MCT油作为快速能量的来源。
1982年,Bach和Babayan[5]综述了饱和脂肪酸在肝脏和肝外组织中的代谢和作用。根据各种脂肪酸的代谢和生理特性,Bach和Babayan提出中链脂肪酸的定义包括C6-C12,长链脂肪酸定义为C14及以上。因此,MCT定义为C8和C10仅在商业上有其起源,而MCFA定义为C6-C12是基于代谢和生理行为。本文将在MCFA和MCT术语中使用Bach和Babayan的定义
椰子油中的脂肪酸和甘油三酯
分析植物油成分有三种常用方法(图2)。最常用的方法是通过化学水解、甲基化和气相色谱分析(方法A)测定植物油的脂肪酸组成。第二种方法需要使用气相色谱或高效液相色谱(HPLC)将油分离成甘油三酯部分,然后通过化学水解(方法B)测定每个甘油三酯(TAG)部分的脂肪酸组成。第三种方法需要使用脂肪酶水解每个TAG部分(方法C)。第三种方法能够对sn-1/sn-3和sn-2位置上的脂肪酸进行区域性鉴定。种方法是最常用的方法,它给出了植物油的脂肪酸谱,但没有给出每种植物油特有的TAG的脂肪酸谱。
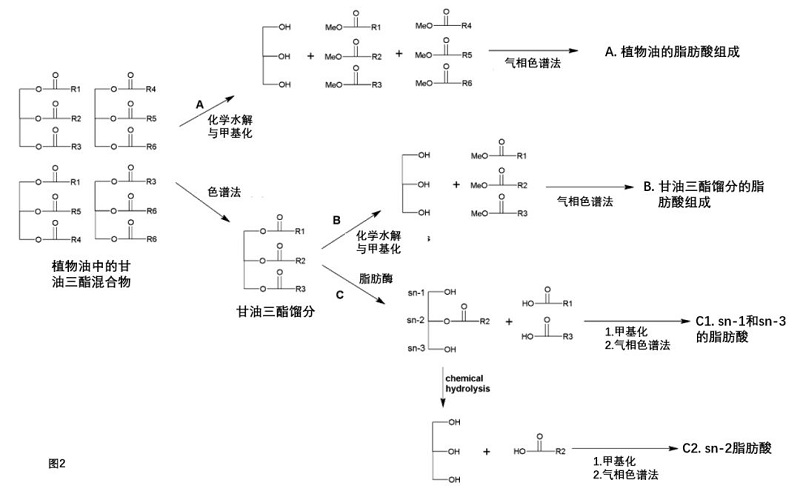
图2. 植物油分析。a化学水解得到植物油的脂肪酸组成。b甘油三酯部分的化学水解产生TAG部分的脂肪酸组成。c利用脂肪酶进行水解,可以识别sn-1/sn-3和sn-2位置的脂肪酸(区域特异性分析)
食品法典命名植物油标准[6]列出了椰子油的脂肪酸组成,椰子油是通过化学水解得到的。Bezard及其同事[7]用气相色谱法分离椰子油中的甘油三酯,然后进行化学水解(方法B),测定了主要三酰甘油酯的脂肪酸组成为:trilaurin(3C12)、caprodilaurin(C10+2C12)、caprolauromyristin(C10+C12+C14)和dilauromyristin(2C12+C14)。Marina和她的同事[8]用高效液相色谱法分析了来自印度尼西亚和马来西亚的12个椰子油样品,使用三酰基甘油标准对其进行了鉴定。收集各馏分,水解后用气相色谱法分析得到各馏的脂肪酸组成。他们测定了主要甘油三酯的脂肪酸组成,并将其定量如下:trilaurin(3C12,23.6%)、capryldilauryl甘油酯(C10+2C12,19.8%)、dicapryllauryl甘油酯(2C10+C12,15.4%)和dilaurylmyristyl 甘油酯(2C12+C14,15.2%)。研究发现,92%的椰子甘油三酯中含有月桂酸,约60%的椰子油中含有MCT。
另一方面,Pham和同事[9]使用对sn-1和sn-3位置有特异性的胰腺脂肪酶分析了来自菲律宾的椰子油。他们确定,54.3%的椰子TAG在sn-1和-3位置含有月桂酸,在sn-2位置含有44.1%。相同位置的癸酸含量分别为4.9%和1.6%(图3)。这意味着至少59.2%(54.3%+4.9%)的椰子TAG的MCFA在sn-1和sn-3位置,至少45.7%(44.1%+1.6%)的MCFA在sn-2位置

图3. 椰子油甘油三酯结构中的月桂酸[10]
新鲜压榨的椰子油(例如,纯椰子油)是甘油三酯、双酸甘油酯、单脂肪酸甘油脂和游离脂肪酸的混合物。使用薄层色谱法(TLC),Pham和同事[10]估计成分约为85%的甘油三酯、7%的双酸甘油酯和3%单脂肪酸甘油脂,而Dayrit和同事[11]使用核磁共振(NMR)光谱法确定非甘油三酯的成分为1.5%的双酸甘油脂、0.01%的α-单脂肪酸甘油脂,以及0.13%的游离脂肪酸。
尽管椰子油和棕榈仁油具有相似的脂肪酸分布(使用方法A),但它们具有明显不同的甘油三酯组成。Bezard[12]分析了棕榈仁油,并报告了其两个主要甘油三酯的不同情况:trilaurin(3C12)和dilauromyristin(2C12+C14)。在对各种植物油的比较研究中,Karupaiah和Sundram[13]确定了椰子和棕榈仁油的区域特异性甘油三酯的分布。他们报告椰子油中最主要的区域特异性甘油三酯种类是(具有指定的sn-位置):1,2,3- trilauryl甘油酯(C12-C12-C12)。
1-己基,2,3-二月桂基甘油酯(C10-C12-C12)和1-己基,2-月桂基,3-肉豆蔻醇甘油酯(C10-C12-C14),而棕榈仁油中的主要甘油三酯是:1,2,3-三月桂基甘油酯(C12-C12-C12),1,2stearyl,3-lauryl 甘油酯(C14-C18-C12),和1,3-油酸,2-月桂基甘油酯(C18:1-C12-C18:1)。对椰子油进行了区域分析,揭示了椰子油的甘油三酯结构和月桂酸的位置分布。月桂酸存在sn-1/sn-3和sn-2位点,对椰子油的消化有重要影响。
椰子油的消化
甘油三酯油的消化涉及物理和酶的过程。椰子油中MCT的比例很大,它与其他植物油(主要是LCT)相比更易溶于水,并且更容易被脂肪酶水解。
舌、胃和胰腺脂肪酶的水解是区域性的,如果MCFA与LCFA相比在TAG中占据sn-1和sn-3位置,则其水解速度更快[14,15]。由于54.3%的椰子TAG含有sn-1或-3位置的月桂酸(如果包括癸酸,则几乎为60%),椰子油被迅速吸收。另一方面,脂肪酶在sn-1和-3位置水解含LCFA和长链多不饱和脂肪酸(PUFA)的TAG的效率较低,导致此类脂肪和油的吸收较慢[17]。
初始水解形成1,2-甘油三酯和游离脂肪酸(图4,步骤1)。第二步水解得到β-单脂肪酸甘油酯和另一种游离脂肪酸。β-单脂肪酸甘油酯经历竞争水解(步骤3)或异构化为α-单脂肪酸甘油酯(步骤4)。因为月桂酸存在于44.1%椰子TAG中的sn-2位置,[18]产生了相对大量的单月桂酸甘油酯。
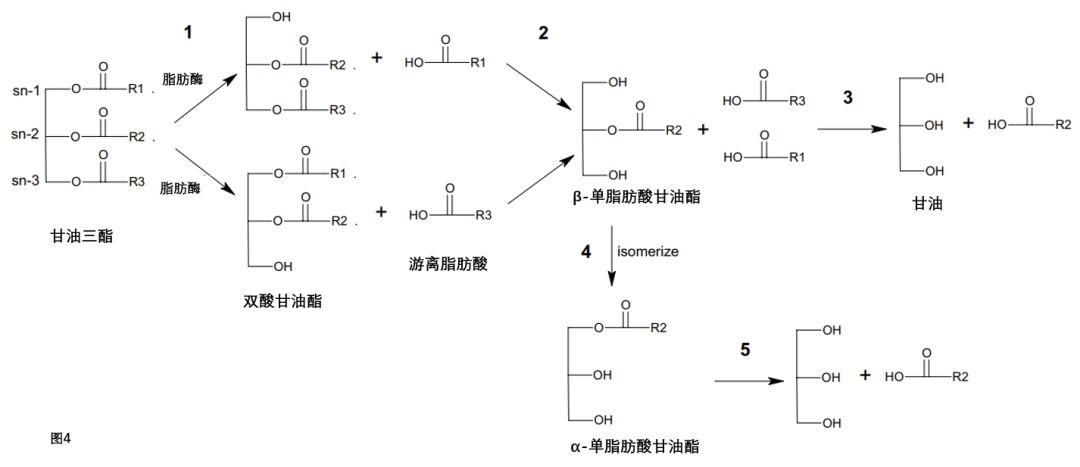
图4. MCT在摄入时经历分步水解
第三个水解步骤产生甘油和从sn-2位置的游离脂肪酸。单脂肪酸甘酯和游离脂肪酸可以被肠细胞吸收,通过特定的载体分子,但也可能通过被动扩散,这取决于它们的链长。由于月桂酸在椰子油中所占比例很高,因此可以假定,摄入椰子油后,单月桂酸甘油酯和月桂酸立即存在于胃中。
Bragdon和Karmen[19]将椰子油喂给人类志愿者,然后将喂食的椰子油的脂肪酸组成与志愿者喂食6小时后收集的乳糜微粒进行比较。乳糜微粒中MCFA(C8-C12)的含量比椰子油低68%,而LCFA(≥C14)明显高于椰子油。据此,他们估计,椰子油中约三分之二的MCFA通过门静脉运输,三分之一进入淋巴并储存在乳糜微粒中。然而,其他研究也表明,MCFA在门静脉和淋巴之间的分布取决于MCT的摄入量与总饮食的比例[20]。也就是说,身体会根据消耗的脂肪和油脂的性质的变化进行调整。
总的来说,研究表明月桂酸具有与癸酸(C10)相似的性质,与棕榈酸(C16)明显不同。此外,含有C6-C12的甘油三酯的代谢特性与含有C14和更长脂肪酸的甘油三酯明显不同。因此,Bach和Babayan提出的MCT中含有月桂酸(C6-C12)的定义是有效的。
月桂酸代谢
月桂酸从甘油三酯释放出来后,通过门静脉直接输送到肝脏,或转化成新的甘油三酯进入淋巴系统。McDonald及其同事[22]通过十二指肠内输注各种脂肪酸,研究了大鼠肠门静脉转运途径。在只注射一种脂肪酸的条件下,他们确定72%的月桂酸、58%的肉豆蔻酸、41%的棕榈酸和28%的硬脂酸绕过淋巴道。也就是说,LCFA比月桂酸更容易进入淋巴道。这项实验还表明,在不存在长链脂肪酸的情况下,月桂酸也可能进入淋巴通路。
研究了月桂酸在大鼠血液中的去向。使用同位素标记的月桂酸和棕榈酸,Goransson[23]观察到,月桂酸比棕榈酸从血液中消失得更快,氧化的速度也更快。他还指出,少量的月桂酸在肝脏中以甘油三酯的形式存在,它没有被纳入磷脂中。然而,他指出,如果老鼠被过度喂养,这些趋势可能不适用。与此结果一致,饮食给药8h后,在大鼠的淋巴脂质中未检测到月桂酸,而检测到肉豆蔻酸和更长的饱和脂肪酸 [24]。这表明,当摄入的月桂酸进入血液时,它会迅速代谢,只有少量以甘油三酯的形式储存在肝脏中。质谱分析来自人类肝组织样本的脂质只检测到LCFA(≥C14)[25]。
在肝脏中,脂肪酸在线粒体中被代谢为能量。根据其链长,脂肪酸可以通过被动扩散或肉碱辅助转运穿过线粒体膜。Gharlid及其同事[26]表明,月桂酸是通过非离子被动扩散快速穿过膜双层的。使用动态核磁共振波谱的测量表明,通过“触发器”机制,MCFA在膜上的扩散速度比LCFA快,碳链延长两个碳会使扩散速度减慢约100倍[27]。与此相一致的是,月桂酸被证明可以在线粒体膜上自由扩散,而长链脂肪酸需要肉碱[28]。因此,月桂酸可以通过物理扩散或在肉碱的帮助下快速运输到线粒体。
月桂酸在肝脏中以多种方式迅速代谢。(图5)β-氧化是柠檬酸循环中脂肪酸代谢产生乙酰辅酶A的主要途径。在人类身上,已经发现四种不同的酰基脱氢酶。确定了每种酶的优选底物为:短链酰基辅酶A脱氢酶(SCAD):C4,C6,C4活性最高;中链酰基辅酶A脱氢酶(MCAD):C4–C14,C6活性最高;长链酰基辅酶A脱氢酶(LCAD):C6–C22,C16活性最高;超长链酰基辅酶a(VLCAD):C12-C22,C16活性最高。其中MCAD和LCAD两种酶对月桂酸有很高的活性。
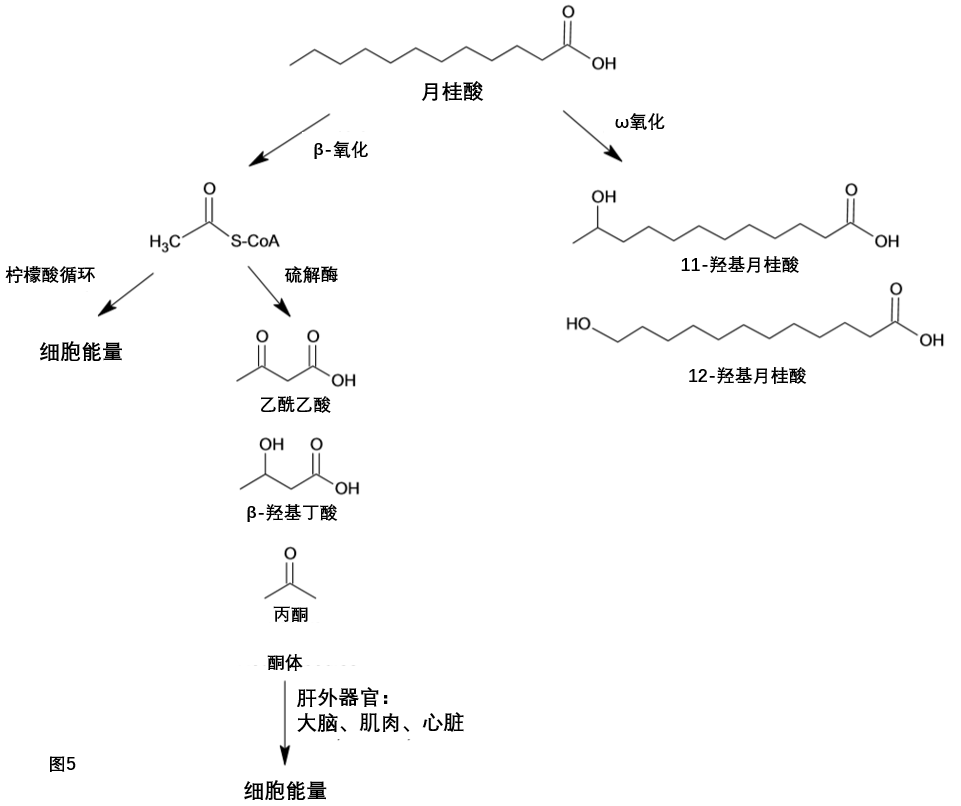
图5. 肝脏中月桂酸的代谢
在肝脏线粒体中,乙酰辅酶A也可以转化为乙酰乙酸,然后转化为β-羟基丁酸和丙酮;这些化合物统称为酮体。虽然肝脏合成酮体,但它几乎没有β-酮酰基辅酶a转移酶,因此不能利用酮体。酮体被输送到其他组织,如大脑、肌肉和心脏,这些组织有酶将酮体转化为乙酰辅酶A作为能量来源
血液中酮体的存在导致胰岛素分泌增加和低血糖。Bach和他的同事给大鼠口服MCT(C6-C12),观察到血浆乳酸、丙酮酸和葡萄糖水平下降,血浆胰岛素水平略有升高。LCT(≥C14)对这些血浆水平没有影响,说明LCT不能有效地生成酮体。
ω-氧化是在正常生理条件下占肝脏脂肪酸氧化总量不到10%的一个次要途径。然而,在饥饿或摄入某些膳食脂肪的情况下,ω-羟化酶的活性可能会增加[31]。月桂酸作为细胞色素P450酶ω氧化的模型底物。许多在肝脏细胞色素P450酶可以催化月桂酸的ω-1和ω-2氧化。细胞色素P450 IVA1(也称为月桂酸羟化酶,LAH)形成11-和12-羟基月桂酸[32]。在人肝脏中,微粒体酶细胞色素P450 4A和细胞色素2E1对月桂酸的ω-1羟基化反应有很好的催化作用[33,34]。月桂酸也可以在肝脏中形成长链脂肪酸。使用放射性标记的1-14C月桂酸,Rioux和同事表明,月桂酸也可以被拉长来产生肉豆蔻酸和棕榈酸[35]。
身体的另一个脂质发挥重要作用的器官是皮肤。皮肤表面的内源性甘油三酯由C16和C18脂肪酸组成,具有单不饱和脂肪酸和支链脂肪酸。月桂酸在皮肤甘油三酯中的含量也较低[36]。月桂酸应用于皮肤外层(角质层)时,由于其分配系数、溶解度参数和构象,对皮肤的亲和力最高。在12小时的皮肤渗透实验中,月桂酸对人体皮肤的渗透性最强[37]。
这些研究表明,月桂酸的代谢和氧化与长链脂肪酸不同。
胆固醇、脂肪堆积和月桂酸
月桂酸本身具有多种调节血清胆固醇水平的作用。如上所述,三酰甘油sn-1和sn-3位置的饱和脂肪酸可导致不同的代谢效应[38]。这意味着膳食脂肪和合成的MCT油可能有不同的生理作用。由于实验设计和喂养时间的不同,关于月桂酸对血清胆固醇的影响一直存在不一致的结论。
1957年,Keys假设饮食中的饱和脂肪会导致高胆固醇血症,而高胆固醇与心脏病有关[39]。Keys将天然饮食中饱和脂肪酸对胆固醇的促进作用缩小到月桂酸、肉豆蔻酸和棕榈酸。然而,由于棕榈酸在美国饮食中比月桂酸和肉豆蔻酸更多,他推测棕榈酸可能是提高美国人血清胆固醇水平的主要原因。
一些饮食实验给出的结果与Keys假设相悖。Hashim和他的同事[41]使用含有C6-C12饱和脂肪酸的MCT制剂进行的一项人类喂养研究表明,血清胆固醇有短暂的上升和下降。他们的研究结果并不支持这样的观点,即高MCFA含量的椰子油会升高血清胆固醇。在对60个饮食脂肪酸影响的对照试验的荟萃分析中,German和Dillard[42]得出结论,月桂酸增加了总胆固醇(TC)和高密度脂蛋白(HDL)。总体效果是TC与HDL的比值降低,这与理想的心血管结果有关。
在文献回顾中,Denke[43]得出结论,从C8到C16的所有饱和脂肪酸都会升高胆固醇,其中C14的作用最强。最近,Tholstrup和他的同事们注意到脂肪酸的位置影响着身体对脂肪的反应。这可能解释了在没有使用天然椰子油的研究中有关月桂酸对健康影响的一些矛盾结论。最后,正如Kaunitz在1970年所指出的,[45]关于饮食中经常食用椰子油的人群的流行病学数据表明,椰子油的食用与冠心病之间没有相关性。例如,对菲律宾波利尼西亚人和比可拉诺斯人作为食用椰子油的自由生活人群的研究表明,椰子油不会对心血管健康产生负面影响,事实上椰子油可能对其他指标有益,比如动脉粥样硬化。
在人类中,已被称为过氧化物酶体增殖物激活受体(PPAR)的核调节蛋白受体,已经被证明可以调节细胞发育和代谢。迄今为止,已经在人体内的各种器官中发现了三个PPAR:PPARα(肝、肾、心脏、肌肉、脂肪组织、其他)、PPARγ(心脏、肌肉、脂肪组织、其他)、PPARβ或PPARδ(大脑、脂肪组织等)。在一些细胞类型中,从C8到C12的MCFA结合PPARγ;然而,在其他细胞中,只有C8和C10 MCFA激活转录。在PPARγ的晶体结构中已经鉴定出了用于MCFA的配体结合口袋。脂质与PPAR的相互作用可以看作是膳食脂肪酸能够调节脂质稳态的分子机制。月桂酸可通过PPARα和γ调节脂肪酸的稳态。
棕色脂肪组织(BAT)是一种专门燃烧脂肪的脂肪细胞。它负责哺乳动物的适应性、非战栗产热。BAT是棕色的,因为有大量含铁的线粒体。产热是指不产生ATP而直接从膳食脂肪中产生体热。BAT的产热能力是由于解偶联蛋白1(UCP1,又称热原蛋白)是线粒体膜上一种独特的BAT特异性转运蛋白,通过电子转运链将质子泵出线粒体后,促进质子的返回。UCP1耗散了驱动细胞ATP合成所需的质子动力,线粒体电化学梯度产生的能量以热的形式释放出来。关于不同脂肪酸作为UCP1激活剂的研究已有很多,但不同文献的结论存在分歧。例如,Shabalina和同事表明MCFA是有效的UCP1激活剂,月桂酸在无细胞系统中激活UCP1[52]。Samartsev及其同事[53]表明,短链脂肪酸的解偶联活性可能高于LCFA,而Fedorenko及其同事更倾向于LCFA的激活。显然,要充分了解这种现象的生热机理以及月桂酸对这种现象的特殊贡献,还需要进行大量的研究。
DeLany和他的同事们设计了一个实验来确定各种膳食脂肪酸导致脂肪形成和肥胖的趋势。他们给体重正常的男性喂食了13c标记的脂肪酸作为他们一周饮食的一部分,并测量呼吸中释放的13CO2。饮食中40%的能量是脂肪,包括以下脂肪酸,按随机顺序摄入:月桂酸、棕榈酸、硬脂酸、油酸、油酸(油酸的反式异构体)、亚油酸和亚麻酸。标记脂肪的累积氧化范围从高达41%的月桂酸到低至13%的硬脂酸,多不饱和脂肪给出中间值。结果表明,在膳食脂肪酸中,月桂酸氧化程度最高,对脂肪积累和肥胖的贡献最小.
月桂酸和单月桂酸甘油酯的抗菌性能
关于月桂酸和单月桂酸甘油酯的体外和体内抗菌性能的研究已经发表了许多报道。在饱和脂肪酸中,月桂酸和单月桂酸甘油酯对革兰氏阳性菌、多种病毒和真菌有很强的抑制作用。
月桂酸、单月桂酸甘油酯和其他衍生物的抗菌活性可分为三种主要机制:1.通过物理化学过程破坏革兰氏阳性菌和脂膜病毒的细胞膜;2.干扰细胞过程,如信号转导和转录,3.稳定人类的细胞膜。这些多重机制的有效性可能是细菌无法进化出抵抗这些化合物作用的能力的原因之一。表1列出了月桂酸和单月桂酸甘油酯抗菌性能的代表性研究结果。

表1. 月桂酸及其衍生物的抗菌活性
早期对脂肪酸抗菌活性的研究发现,月桂酸是饱和脂肪酸中最活跃的[56,57];系统研究了脂肪酸、单脂肪酸甘油酯和双酸甘油酯表明的体外抗菌活性,月桂酸是对抗革兰氏阳性菌最有效的脂肪酸[58],而α-单月桂酸甘油酯比月桂酸更有效[59]。在月桂酸存在下,感染性水疱性口炎病毒的产生受到剂量依赖性和可逆性的抑制:去除月桂酸后,抗病毒作用消失。此外,饱和脂肪酸的链长被证明是至关重要的,因为链短或链长的脂肪酸效果较差或没有抗病毒活性[60]。但是,月桂酸的活性被Mg2+和Ca2+离子降低,而较低的pH值通常会提高其活性。这些观察结果表明,月桂酸的吸收受酸和细菌细胞表面的理化性质的控制[61,62]。在牙科护理中,月桂酸可以减少牙菌斑的形成并抑制羟基磷灰石的溶解]。
除了抗菌活性外,单月桂酸甘油酯还被证明能有效阻止或延迟致病性革兰氏阳性细菌产生外毒素。单月桂酸甘油酯被证明是在转录水平,抑制葡萄球菌毒素和其他外蛋白合成的机制。此外,研究还表明,单月桂酸甘油酯可以通过干扰信号转导阻断β-内酰胺酶的诱导。单月桂酸甘油酯抑制金黄色葡萄球菌毒力因子的表达,抑制粪肠球菌对万古霉素的耐药性。提示单月桂酸甘油酯通过阻断信号转导而起作用。
在一系列脂肪酸和单脂肪酸甘油酯中,月桂酸和单月桂酸甘油酯对幽门螺杆菌具有很强的抑制作用。此外,幽门螺杆菌对单月桂酸甘油酯杀菌活性的自发抗性较低[68]。白色念珠菌对几种脂肪酸及其α-单脂肪酸甘油酯的敏感性表明,在较低浓度和较长的培养时间后,C10作用最快,而C12最活跃。此外,这些脂肪酸对皮肤和粘膜没有毒性,是外用的理想选择。通过比较饱和脂肪酸C10-C18对胡宁病毒(JUNV)感染的抑制作用,发现月桂酸是最有效的抑制剂。从机理研究中可以得出结论,月桂酸抑制了JUNV复制周期中的一个晚期成熟阶段。
月桂酸及其衍生物(包括单月桂酸甘油酯)的作用机理引起了人们的极大兴趣。经月桂酸处理的产气荚膜梭菌的透射电镜图像显示细胞内外膜分离和胞质紊乱。此外,月桂酸被发现是C8-C14系列中最活跃的饱和脂肪酸[71]。在一项比较饱和链与阳离子、阴离子和非离子基团的研究中,卡巴拉得出结论:一般来说,阳离子物种(如含铵基团)比阴离子和非离子物种更为活跃,链长为10-16个碳原子。然而,阳离子物种的毒性高于阴离子物种[72]。非离子单脂肪酸甘油酯表现出活性,特别是当酯基为月桂基时。
其他研究人员认为,单月桂酸甘油酯的独特活性可能是由于其化合物本身及其水解产物月桂酸的抗菌作用。许多月桂酸衍生物的抗菌活性,包括单月桂酸甘油酯,支持这一假设(见表2;图6)。α-单月桂酸甘油酯(MLG-1)和α-单月桂基甘油醚(MLE-1)(α-单月桂酸甘油酯的醚类似物)的比较是说明性的。在体内,MLG-1抑制金黄色葡萄球菌的生长,但被金黄色葡萄球菌脂肪酶降解。另一方面,MLE-1在体内对金黄色葡萄球菌脂肪酶稳定,但效果不如MLG-1[74]。MLG-1和MLE-1的关键区别在于MLG-1在水解时释放另一种抗菌化合物C12的能力。一旦C12被释放,它就被缓慢代谢,这延长了它的作用[75]。因此,MLG-1是一种比MLE-1更有效的抗葡萄球菌局部抗感染药物。
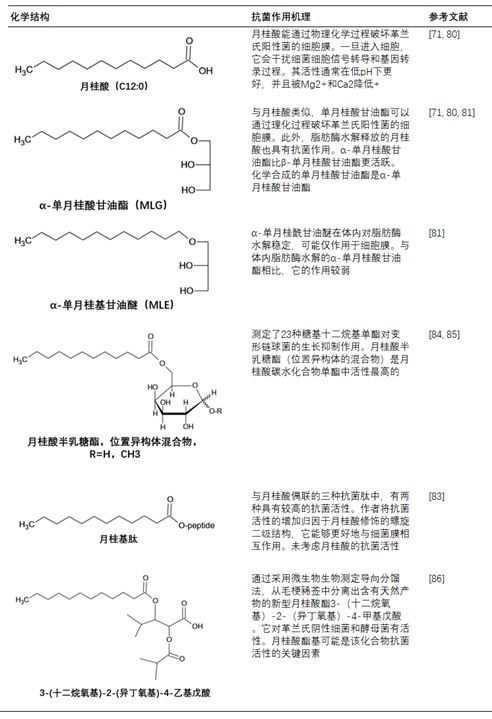
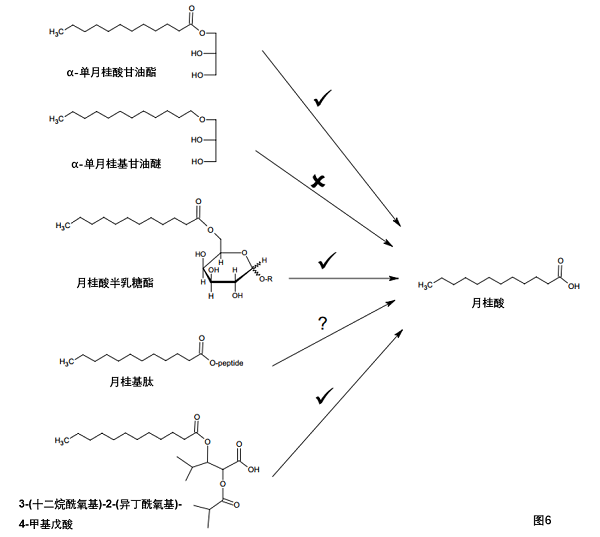
表2. 月桂酸及其衍生物的化学结构与抗菌活性的关系,与脂肪酶或其他酶的的反应见图6
含有月桂基酯的各种化合物已被证明是抗菌的。与月桂酸偶联的肽具有较高的抗菌活性。抗菌活性的增加归因于由于月桂酸使肽能够更好地与细菌膜相互作用,而导致的螺旋结构的变化。抗菌活性的增加可能是由于月桂酸本身引起的,这一点未被考虑。糖基月桂酸衍生物作为食品致病菌的生长抑制物具有很强的应用前景。例如,甲基α-d-甘露吡喃苷的月桂酸酯成功地控制了李斯特菌和其他革兰氏阳性病原体,而半乳糖和果糖月桂酸酯对变形链球菌的细胞生长具有强烈的抑制作用。
近年来,利用金黄色葡萄球菌(S.aureus)对传统药用植物毛梗豨莶草(菊科植物)的地上部分进行生物测定指导下的分离,分离出了一种新型的含月桂基酯的天然产物3-(十二烷氧基)2-(异丁氧基)-4-甲基戊酸。该化合物对革兰氏阳性菌(枯草芽孢杆菌、粪大肠杆菌、痤疮丙酸杆菌、表皮芽孢杆菌、猪链球菌亚种 凝集素,无乳链球菌和热原菌),革兰氏阴性菌(大肠杆菌和铜绿假单胞菌)和酵母菌种类(无脂酵母菌和新生酵母菌)。与单月桂酸甘油酯的情况一样,月桂酸酯基团可能是该化合物抗菌活性的关键。
月桂酸和单月桂酸甘油酯的一个重要特征是没有人体毒性。母乳含有约3.5–4.5%的脂肪,主要是甘油三酯,其中约4–6%是月桂酸。当母乳被消化时,释放的单脂肪酸甘油酯和游离脂肪酸会强烈抑制包膜病毒、一些细菌和原生动物。母乳中最强的抗菌剂是单月桂酸甘油酯。对葡萄球菌属、棒状杆菌属、芽孢杆菌属、李斯特菌属和链球菌属的革兰氏阳性菌进行了MCFA及其1-单脂肪酸甘油酯的抗菌活性研究。α-单脂肪酸甘油酯比相应的脂肪酸更具活性;仅C12是弱活性的。单葵酸甘油酯与单月桂酸甘油酯之间存在协同作用。这些信息被用来了解人类初乳的抗菌特性,并设计婴儿配方奶[82]。单月桂酸甘油酯是唯一一种被发现用作食品防腐剂的单脂肪酸甘油酯。单月桂酸甘油酯已被证明可以抵御食源性病原体并抑制牛奶中的大肠杆菌O157:H7。月桂酸及其衍生物在治疗动物体内的微生物感染方面也有重要应用,如奶牛中的金黄色葡萄球菌和家禽、仔猪和兔子中的产气荚膜梭菌。
单月桂酸甘油酯展示了涉及哺乳动物细胞膜稳定的第三种作用模式。观察到单月桂酸甘油酯插入人细胞膜,从而阻止信号转导机制,如宿主细胞受体信号系统已被外毒素篡夺。它还能够稳定红细胞,抵抗溶解作用。
1977年,卡巴拉注册了合成单月桂酸甘油酯,商标是Lauricidin™ ,作为食品级杀微生物剂或抑菌剂,申请了批商业用途的专利之一[89]。从那时起,在全世界范围内已经有超过100项关于使用单月桂酸甘油酯的专利申请,涉及食品和非食品的各种应用,如医疗程序、消毒和消毒剂、抗菌聚合物成分、动物饲料补充剂等。
总结和结论
月桂酸约占椰子油中脂肪酸的一半,详细的研究表明,月桂酸对椰子油的许多特性起着重要作用。椰子油的甘油三酯结构使椰子油比其他以长链脂肪酸(LCFA)为主的植物油能更快地被消化。在正常饮食中,摄入的大部分月桂酸通过门静脉直接输送到肝脏。虽然乳糜微粒中可能发现少量的月桂酸以甘油三酯的形式存在,但在磷脂中没有发现。与这些特性一致,椰子油尚未被证明会导致心血管疾病和动脉粥样硬化。
月桂酸通过被动扩散迅速地通过线粒体膜转运,不需要肉碱辅助转运。月桂酸在肝脏中以多种方式迅速代谢。两种酰基辅酶A脱氢酶能迅速氧化月桂酸。月桂酸可代谢为酮体,酮体是体内肝外器官如大脑、心脏、肌肉的重要能量来源。在所有脂肪酸中,月桂酸对脂肪积累的贡献最小。月桂酸的这些性质与椰子油是一种非育肥能源的观察结果一致。
在所有饱和脂肪酸中,月桂酸对革兰氏阳性菌、某些病毒和真菌的抗菌活性最强。这些化合物具有独特的之处在于它们能够避免微生物产生耐药性,这可能是由于它们的多种作用,包括破坏细胞壁和干扰细胞信号和转录。事实上,在所有单脂肪酸甘油酯中,单月桂酸甘油酯具有最强的抗菌活性。它可以作为一个完整的化合物,也可以在水解后释放月桂酸。基于单月桂酸甘油酯的专利和商业产品很多。
椰子油有许多有益的作用。这篇关于月桂酸的综述为椰子油的许多有益作用提供了理论支持。最后,由于MCFA(C6-C12)表现出与LCFA(≥C14)足够不同的代谢和生理特性,在使用术语“饱和脂肪酸”时应指定链长。