细胞骨架(cytoskeleton)是指真核细胞中的蛋白纤维网络结构。发现较晚,主要是因为一般电镜制样采用低温(0-4℃)固定,而细胞骨架会在低温下解聚。直到20世纪60年代后,采用戊二醛常温固定,才逐渐认识到细胞骨架的客观存在。
细胞骨架不仅在维持细胞形态,承受外力、保持细胞内部结构的有序性方面起重要作用,而且还参与许多重要的生命活动(图1),如:在细胞分裂中细胞骨架牵引染色体分离,在细胞物质运输中,各类小泡和细胞器可沿着细胞骨架定向转运;在肌肉细胞中,细胞骨架和它的结合蛋白组成动力系统;在白细胞的迁移、精子的游动、神经细胞轴突和树突的伸展等方面都与细胞骨架有关。另外,在植物细胞中细胞骨架指导细胞壁的合成。
细胞骨架由微丝(microfilament)、微管(microtubule)和中间纤维(intemediate filament)构成。微丝确定细胞表面特征,使细胞能够运动和收缩。微管确定膜性细胞器(membrane-enclosed organelle)的位置和作为膜泡运输的导轨。中间纤维使细胞具有张力和抗剪切力。
微丝、微管和中间纤维位于细胞质中,又称胞质骨架,它们均由单体蛋白以较弱的非共价键结合在一起,构成纤维型多聚体,很容易进行组装和去组装,这正是实现其功能所必需的特点。
广义的细胞骨架还包括核骨架(nucleoskeleton)、核纤层(nuclear lamina)和细胞外基质(extracellular matrix),形成贯穿于细胞核、细胞质、细胞外的一体化网络结构。
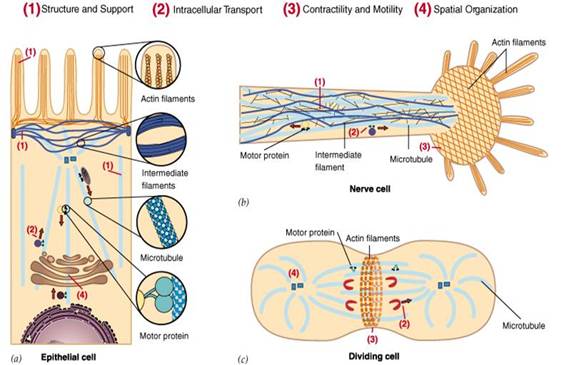
图1为细胞骨架的主要功能
细胞骨架是细胞功能的结构基础,在外界刺激或在细胞分裂、分化过程中,细胞形态发生相应的变化,这和信号传导调控细胞骨架的动态学变化分不开的。细胞骨架作为真核细胞中的动态蛋白纤维网结构,由微丝(microfilaments, MF)、微管(microtubules, MT)和中间纤维构成(intermediate filaments, IF)。MF是由肌动蛋白聚合而成的多聚体, 广泛存在于真核细胞应力纤维、黏着斑、伪足和收缩环中, 其结构组成单元球状肌动蛋白(globular actin, G-actin)在ATP和多种微丝相关蛋白的作用下, 通过与丝状肌动蛋白(filamentous actin, F-actin)不断组装和解聚的转换, 参与调控细胞形态改变、黏附、迁移和胞质分裂等行为。MT由α-tubulin和β-tubulin聚合组成, 在微管相关蛋白的作用下组装形成中空管状结构, 在细胞内物质运输与胞质分裂中扮演重要角色。MF和MT细胞骨架相互联系、相互依赖, 共同参与调控细胞行为(Bailly M,2002; Miranti C.K,2002)。
细胞迁移是恶性肿瘤侵袭和转移中的关键步骤,细胞骨架和其结合蛋白是细胞迁移过程的物质基础,细胞定向运动需要细胞骨架的参与,尤其是由肌动蛋白组成的微丝骨架。MF和MT不仅在稳定细胞形态、承受外力刺激、维持细胞内部结构有序性中起重要作用, 而且还参与调控细胞迁移、黏附、分裂以及细胞内信号传递。其中, cAMP介导的信号通路与细胞骨架之间存在的互动效应与细胞增殖、凋亡、黏附、迁移等细胞行为密切相关, cAMP依赖的蛋白激酶A(PKA)作为cAMP依赖的主要靶向蛋白, 参与了这些过程。PKA通过磷酸化作用激活RhoA, RhoA激活后除了进一步激活下游通路外, 还增强与Rho二磷酸鸟苷解离抑制因子(RhoGDI) 的结合能力, 从而反向抑制RhoA活性(Mehlen, P,2006)。
整合素是细胞表面一类重要的兼具粘附和信号转导功能的受体,它是由α和β亚基通过非共价键组成的异二聚体。细胞外基质蛋白,如纤粘连蛋白、层粘连蛋白和胶原等是其主要配体;其次还有一些细胞表面分子,可溶性蛋白等也可与整合素结合。通过胞外段与胞内段与细胞骨架、信号转导分子和其它一些蛋白相结合,整合素象是一座桥梁介导了细胞内外之间的双向信息传递。整合素可以与细胞骨架之间的双向信号转导,(一)从细胞骨架到整合素,整合素在没有与结合时是分散在细胞表面的,而且不与肌动蛋白细胞骨架相连,一旦与ECM中的配体结合,则通过其胞内域与细胞骨架发生联系,并发生簇集。(二)从整合素到细胞骨架,整合素可反馈调节细胞骨架的组装,整合素介导的粘附自身就可以激活Rho家族小G蛋白(Mitra S.K,2005;Larsen M,2003; Turner C.E,2000)。